第四章 视觉
欲求得合适的答案,需先问出恰当、关键的问题。
视杆细胞,负责夜间视觉,数目更多。集中在中心凹。
视锥细胞,负责高度敏锐视觉和色觉。
-
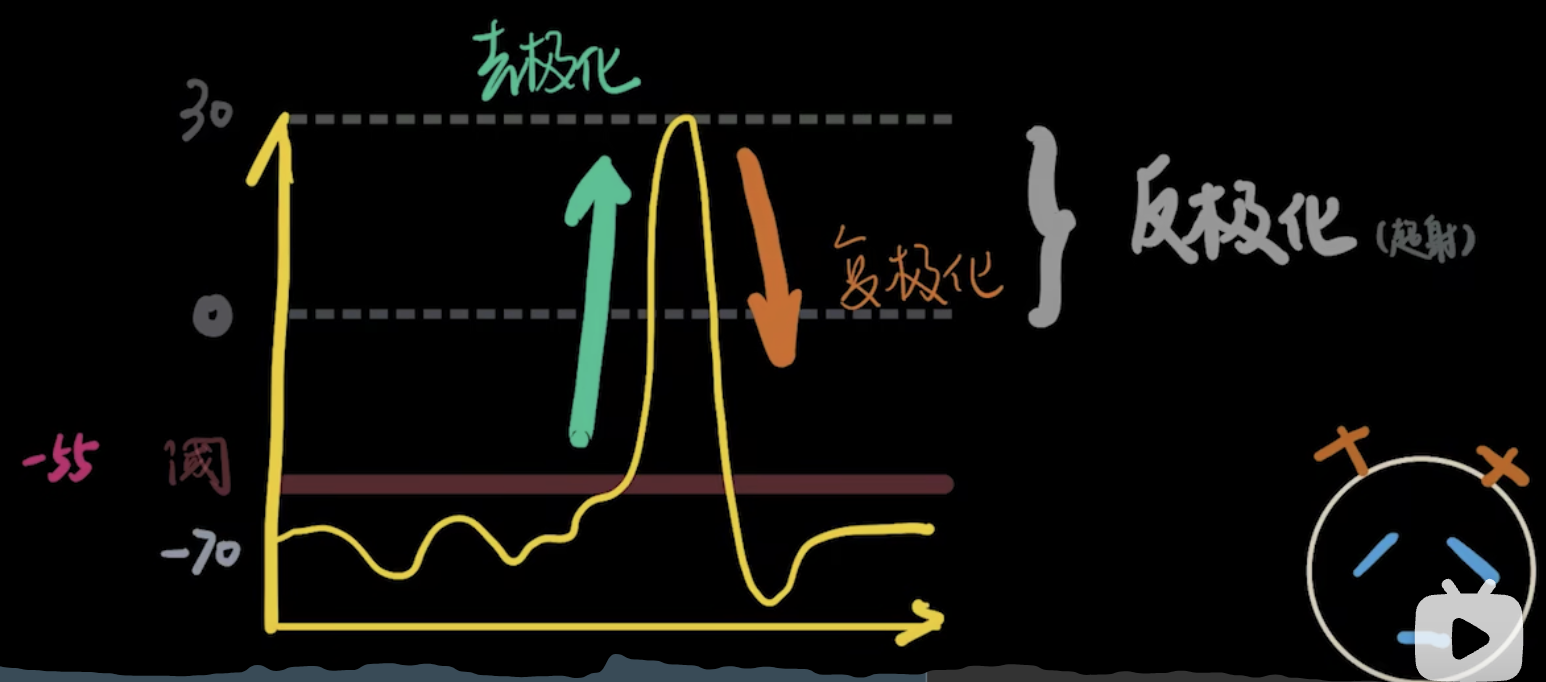
去极化,复极化和反极化
-
超极化 细胞进一步拉大内外的电位差
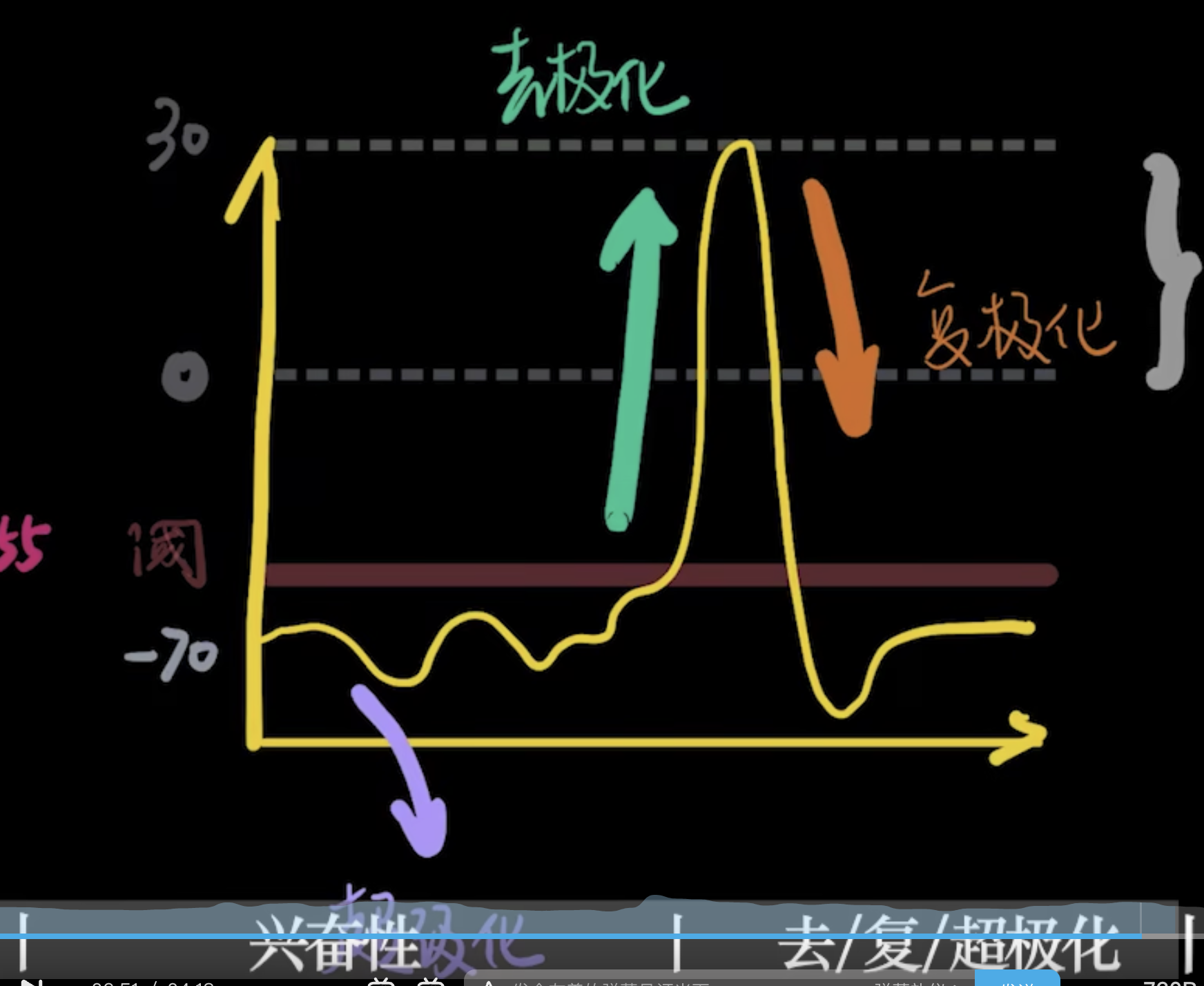
电压门控Na+通道关闭比较及时,但是电压门控K+通道关闭很缓慢,超过了-70mv仍旧外流。但是有钠钾泵的存在,可以将电位恢复到静息电位。
激活函数可以引入一个当前值,模仿去极化的,复极化等过程。针对视频处理可能有用,但是针对图片处理可能还需要在做出调整。或者加入缩放和偏置。
脊椎动物的视杆细胞和视锥细胞是利用超极化来报告感觉刺激的。
向视杆细胞外段注射cGMP可以模拟黑暗环境导致视杆细胞去极化。
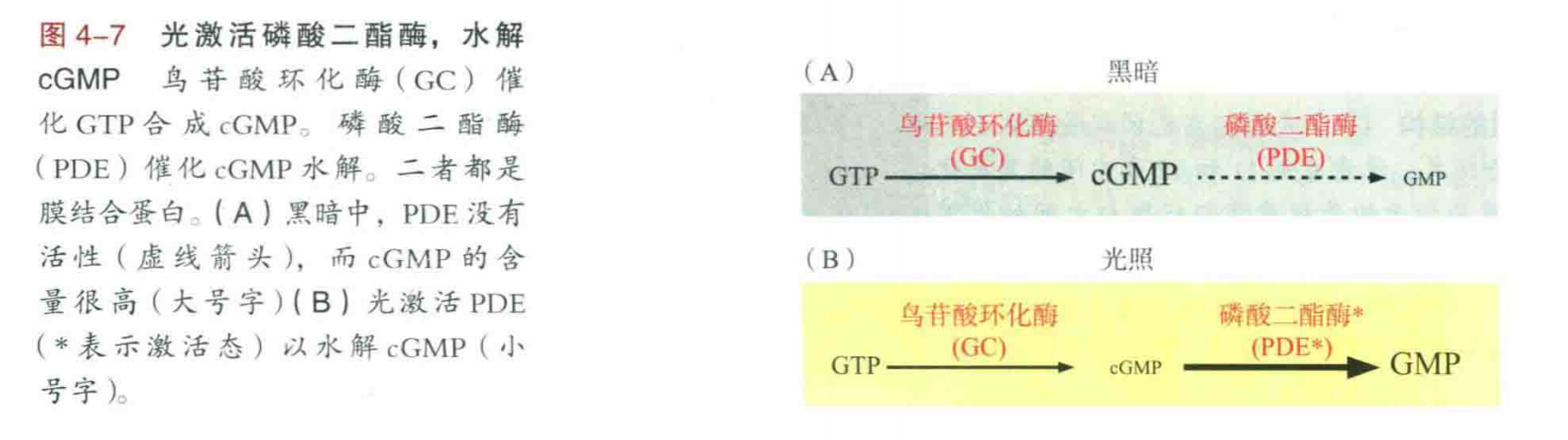
光信号传导的级联过程,一个光激活的视紫红质可催化超过20个转导蛋白分子,每个转导蛋白激活一个PDE,最终导致每秒钟数以万计的cGMP水解。生化级联反应中的信号放大作用解释了单个光子如何触发了cGMP水平的大幅度下降。cGMP的浓度下降导致了cGMP门控阳离子通道关闭,削弱了稳定的阳离子内流。这使得视杆细胞超极化,从而减少了视杆和视锥细胞所使用的神经递质的释放。
一个最优化的感觉系统还需要综合其他的一些特性,例如响应速度,动态范围,选择性和可靠性等。
复原:被光激活的感光细胞重新回到暗态的过程。复原作用保证一个视觉刺激只产生一个瞬时信号,且同一个感光细胞在数秒内能准备好接受下一个视觉刺激。
所有光信号传导级联过程中被激活的视杆细胞的组分都必须迅速回到暗态,暗态即视杆细胞准备好进行光接收和转导的状态。
激活函数设计成一个有限状态自动机?
复原过程中,cGMP的水平必须上升(笔者注:意味着回到暗态),才能重新打开CNG通道,从而使视杆细胞去极化。复原过程的细节没看懂,只知道:光照降低了cGMP水平,关闭了cGMP门控通道,但会触发一个负反馈环路,导致cGMP水平的提高,又打开了cGMP门控通道并恢复了暗态。
适应使得视觉系统能在大范围的光照水平中检测到反差。对目标的检测需要做到将目标物与其背景区分开来。目标检测的一个基本手段就是利用亮度反差。
我们的视觉系统有着非常大的动态范围(定义为光强最大值和最小值的比),使得我们既能在接近黑暗的状况下检测出单个光子,也能在10^11倍范围内的环境光照水平下从背景中分辨目标。
视杆细胞负责的是动态范围中光强较低的部分,能在10^4倍的范围内的环境光照水平下辨别出亮度反差。
适应:感光细胞可以根据背景的光照水平而调节自身灵敏度的机制。
韦伯定律:尽管不能对所有的刺激形式都适用,但是该定律描述了恰可分辨两个感觉刺激的差值与刺激的量级成比例的情况。比如人类能区别100g和105g的差别,但是无法区分1000g和1005g的差别。
光适应性:当背景的亮度升高时,感光细胞对于同等的光强的敏感度降低,为了在更高的亮度的背景下达到相同的超极化的量,需要一个更强的刺激才行。
视锥细胞密集分布在中央凹,负责高精度视觉。视锥细胞和视杆细胞的功能互补:负责高亮度的视觉,高空间精度的视觉,范围更广的运动视觉和颜色视觉。事实上,对于人类的日常生活而言,视锥细胞要比视杆细胞重要得多。
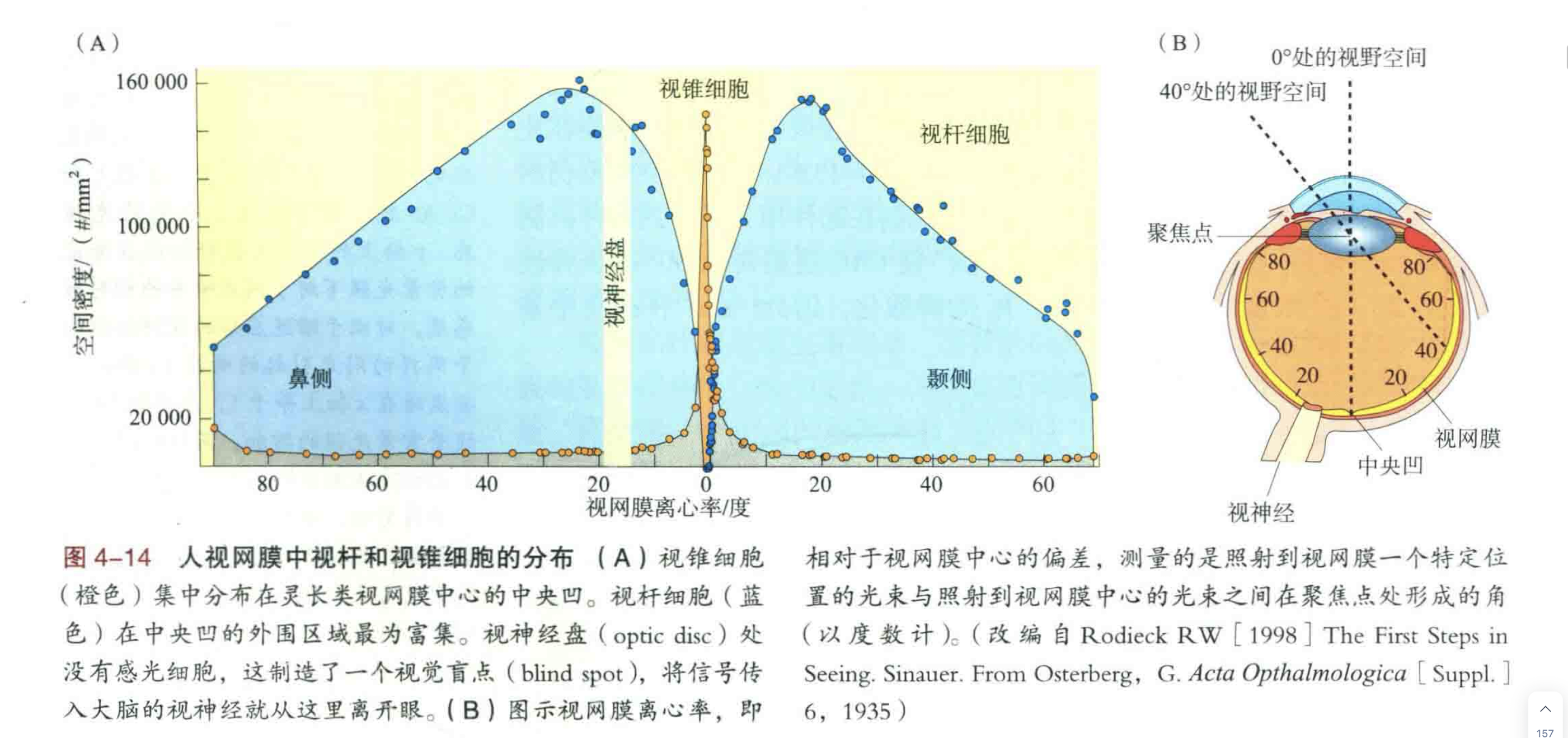
灵长类的高精度视觉依赖于集中在中央凹处高密度的视锥细胞。相比于感光细胞分布较为稀疏的外周视网膜,中央凹处相互紧邻的视锥细胞能够分辨出更小的光点。
灵长类的视网膜处理信息所用的大部分能量都消耗在中央凹上,尽管中央凹只占视网膜总面积的1%,但从中央凹到大脑的信息传递却占据了约一半的视网膜节细胞的轴突。
视杆细胞和视锥细胞的比较:
两者在感光和速度上具有重要差异。
视锥比视杆的感光性弱得多。
但是视锥细胞的光响应要比视杆细胞快得多。
另外,视锥细胞的响应比视杆细胞更迅速回到基线,所以视锥可以对更快速的重复光刺激产生响应,因此拥有更高的时间分辨率。这对于运动的检测十分重要。
这些差异,反映了两种细胞在光信号转导级联过程和复原机制上的不同。
视锥细胞比视杆细胞能适应更宽范围的光强度。视锥负责在10^11倍范围的环境光强分辨物体。
人类有三种类型的视锥细胞:S视锥细胞,M视锥细胞,L视锥细胞。
S视锥细胞要比M、L稀疏很多,仅占中央凹视锥细胞总数不到5%。这解释了为何我们队蓝光的空间分辨率不高。
事实上,由于不同波长的光在通过我们眼前部的晶状体时所受到的折射不同,它们无法以相同的锐度聚焦在视网膜上,这一性质叫做色差。我们演化出了对长波光更敏锐的视觉,所以即使蓝色视锥细胞密度更高一些,我们也不会获得比蓝光更好的分辨率。
M和L视锥细胞的比例在不同个体有所差异,且M和L视锥细胞的空间分布是随机的。
外界光信号在视锥细胞和视杆细胞上产生的信息在视网膜是上如何加工的呢?
捕捉光信号的过程中,视网膜上的感光细胞会将外界的光刺激的强度转化为随时间变化的二维阵列信号。动物需要从外界光信号中提取有效的特征信息,如物体的运动信息,深度信息,颜色信息和大小信息。
以人为例,人的视觉神经系统包含大约1亿个感光受体,但仅有约100万视网膜节细胞将外界的光信号传输到大脑。因此在视觉处理过程中,我们的视觉系统会选择保留有行为意义的视觉信号,而尽可能多地剔除掉不相关的信息。
感受野:感觉生理学各出的概念,这个概念来源于之前对躯体感觉的研究。躯体感觉通路中某个神经元会因为刺激身体某些部分而兴奋,所有能使该神经元兴奋的身体部位称作该神经元的感受野。
视觉系统中某神经元的感受野指的是当它兴奋时,光刺激照射在视野内的部分。
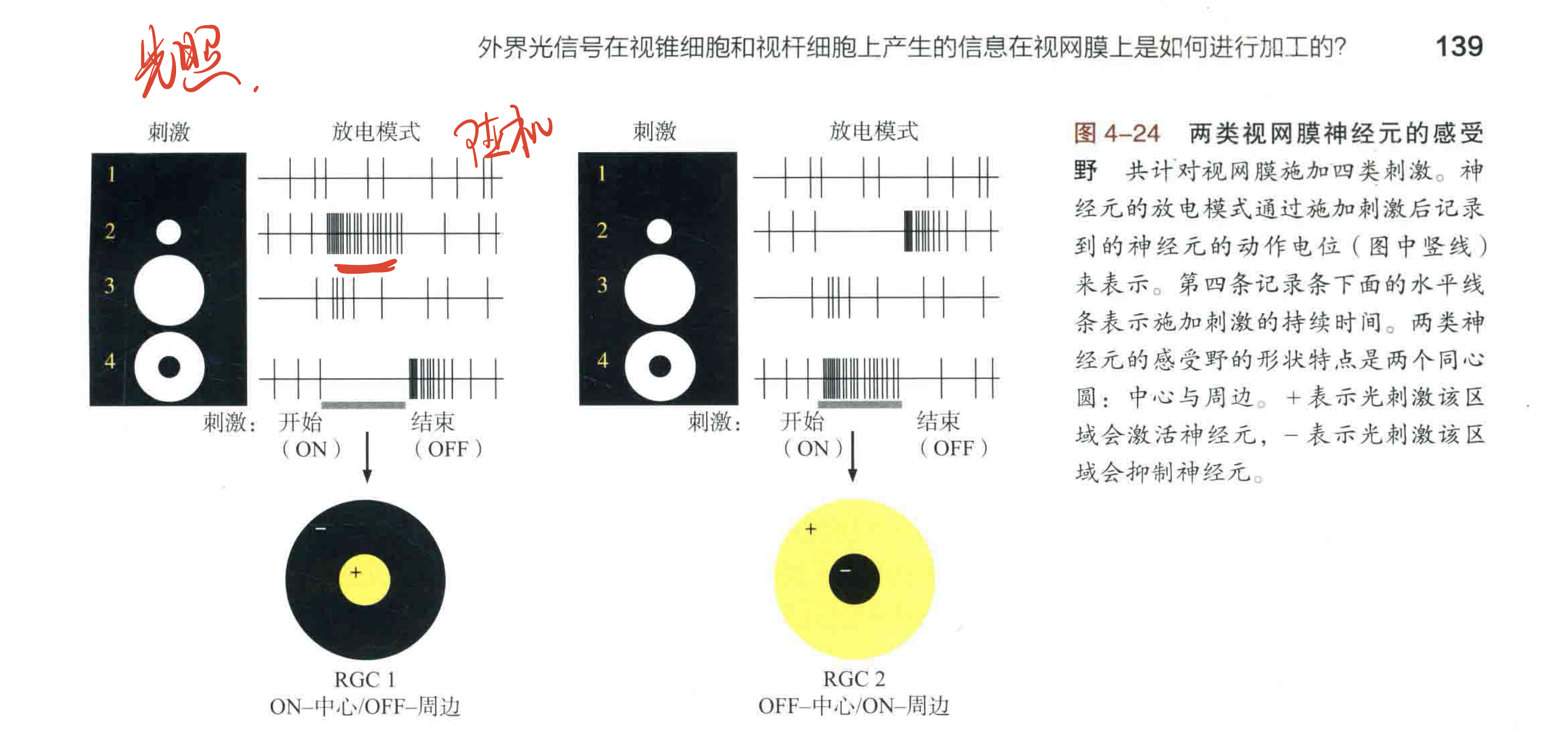
视觉处理的上游-将视网膜电信号传输到大脑的视网膜节细胞
光刺激是中心亮的时候,神经元1发放的动作电位最多,随着明亮区域增大,动作电位发放频率降低。当明亮区域增大到一定值时,再增大其面积不会改变神经元的兴奋性。这是因为光照再中心区域会提高神经元的兴奋性,但光照再外周区域会降低神经元的兴奋性。
给出的第四类刺激,中央直接不给刺激,只在四周施加光照,产出的放电模式可以看出,神经元几乎不再产生动作电位,兴奋性降低。
然后下图右侧就是神经元2,感受野性质和神经元1完全相反。只有外周施加光照才能最大程度刺激神经元2的兴奋,中央施加光照反而会抑制神经元2的兴奋性。
还有一个有趣的现象,下面两种神经元都具有的特性,就是在抑制的情况下(如刺激4对神经元1的抑制或者刺激2对神经元2的抑制)撤除光照刺激之后,动作电位发放频率都有瞬时的增加,虽然会很快恢复到静息的发放频率。
神经元1被称为 中心激活-外周抑制型;神经元2被称为 中心抑制-外周激活型。
这种视网膜神经元中心-周边感受野的性质对于我们理解视觉信号处理有重要启发:视网膜神经元细胞不仅仅对光信号做出反应,还可以进行小面积视网膜上的明暗对比,从而确定光信号的空间信息。